
獲得遺伝子(哺乳類特異的遺伝子)から哺乳類らしさにせまる


獲得遺伝子(哺乳類特異的遺伝子)から哺乳類らしさにせまる
哺乳類特異的なエピジェネティクス機構の ゲノムインプリンティング の研究から、私たちは胎盤形成に必須の PEG10 遺伝子 を発見しました。
この遺伝子はウイルスによく似たDNA配列を持つ 外来DNA由来の獲得遺伝子で、胎生の哺乳類にのみ存在します。現在では、お母さんが赤ちゃんを
産み育てることは、哺乳類にとって自然なことであり “必然” のことに思われます。しかし、胎生の起源 を哺乳類のゲノムの中に探求していくと
幾つもの 哺乳類特異的獲得遺伝子 が “哺乳類らしさ” の形成に重要な役割を果たしたことがわかりました。 ヒトやマウスを含むグループには、
PEG10 の他にもこのような哺乳類特異的な獲得遺伝子が多数存在し、胎盤だけではなく脳でも重要な機能を果たしています。
“偶然”飛び込んできた外来のDNAのうちのごく一部が、哺乳類のゲノムとして利用され、有益な遺伝子として選択されることで、現在の“必然”
としての日常があると言えるでしょう。私たちの研究は、獲得遺伝子が哺乳類レベルの進化に関与した具体例を示したものであると考えています。
ゲノムインプリンティング は哺乳類特異的な遺伝子発現機構です。私たちは、このゲノムインプリンティングを切り口として、哺乳類の特徴である生殖様式(胎生)がどのように成立したのかという問題に迫り、胎盤形成に必須の PEG10 遺伝子 を発見しました。
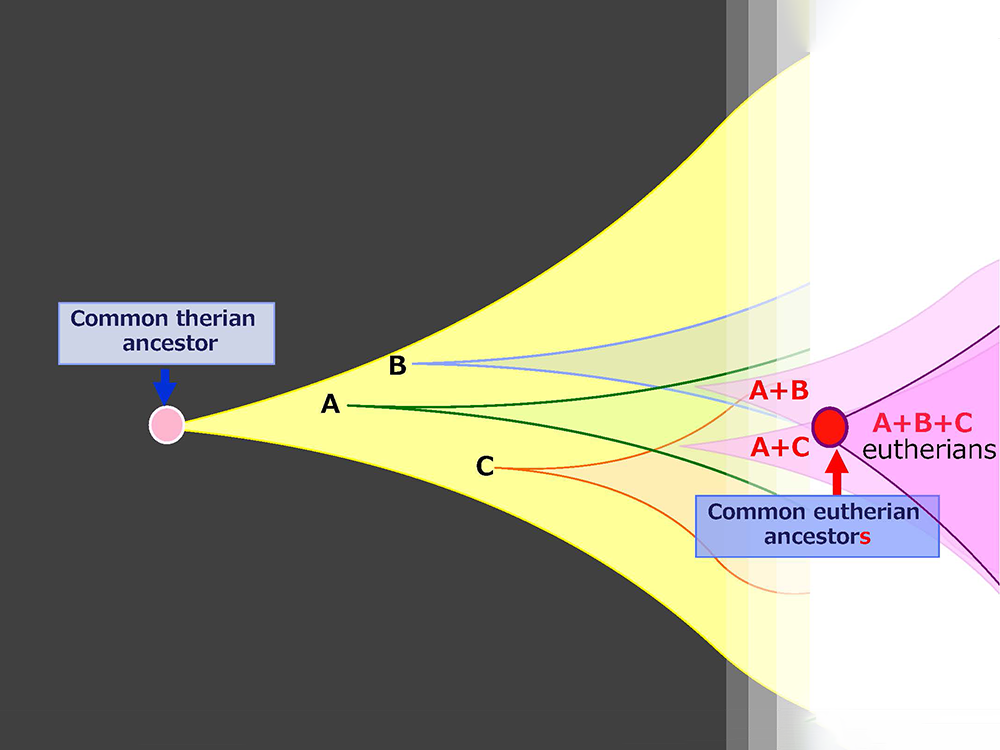
1990年代に、ゲノムインプリンティングに関わるインプリント遺伝子の網羅的探索を始めるにあたって、インプリント遺伝子には哺乳類になって獲得した遺伝子が含まれ、その新しい遺伝子が哺乳類の特徴を生み出している可能性があると考えていました。そして、新しい遺伝子のリソースには感染したウイルスゲノムの断片を想定していました(図1)。
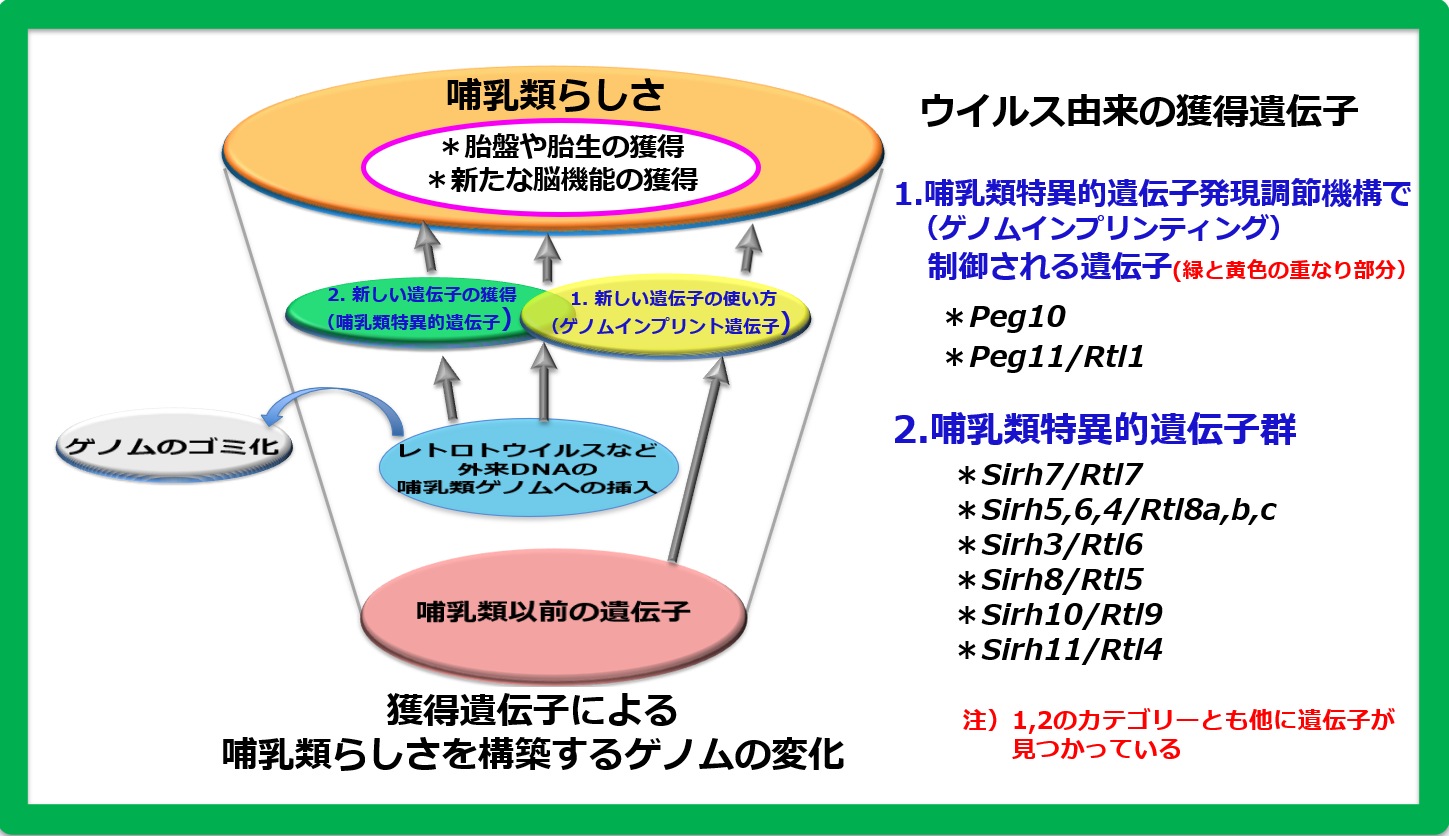
Peg10遺伝子はウイルスによく似たDNA配列を持つ 外来DNA由来の獲得遺伝子で、胎生の哺乳類にのみ存在します。現在では、お母さんが赤ちゃんを産み育てることは、哺乳類にとって自然なことであり “必然” のことに思われます。しかし、この胎生という生殖機構の成立には、ウイルスの感染からの遺伝子獲得という “偶然” の出来事が大きな役割を果たしていたことがわかったのです。胎生の起源 を哺乳類のゲノムの中に探求していくと、ゲノムインプリンティングとは無関係のさらに幾つもの 哺乳類特異的獲得遺伝子が胎盤形成に関係していることや、獲得遺伝子が哺乳類の特徴である発達した脳機能にも大きく関係していること、言い換えれば“哺乳類らしさ” の形成に重要な役割を果たしたことがわかりました。
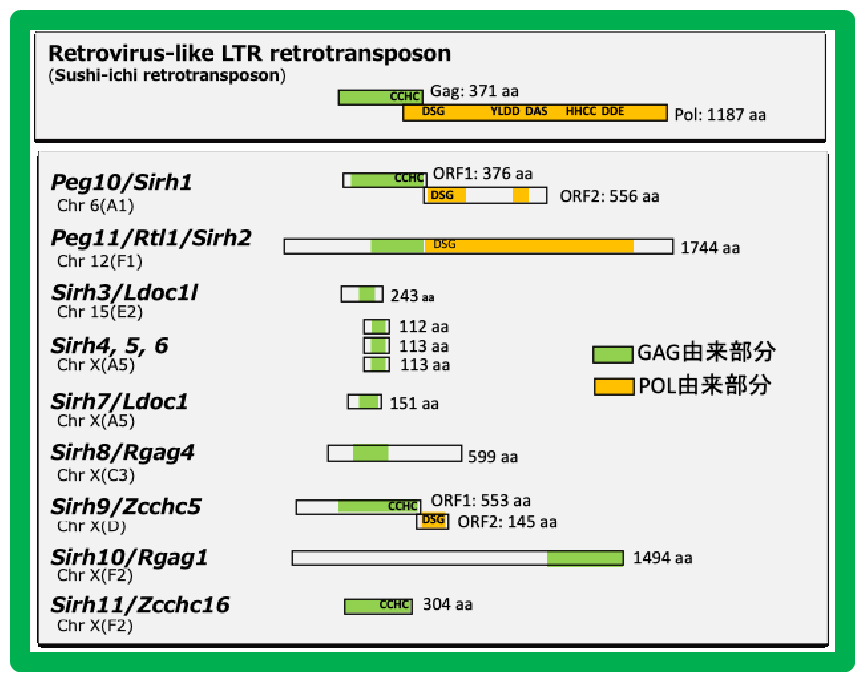
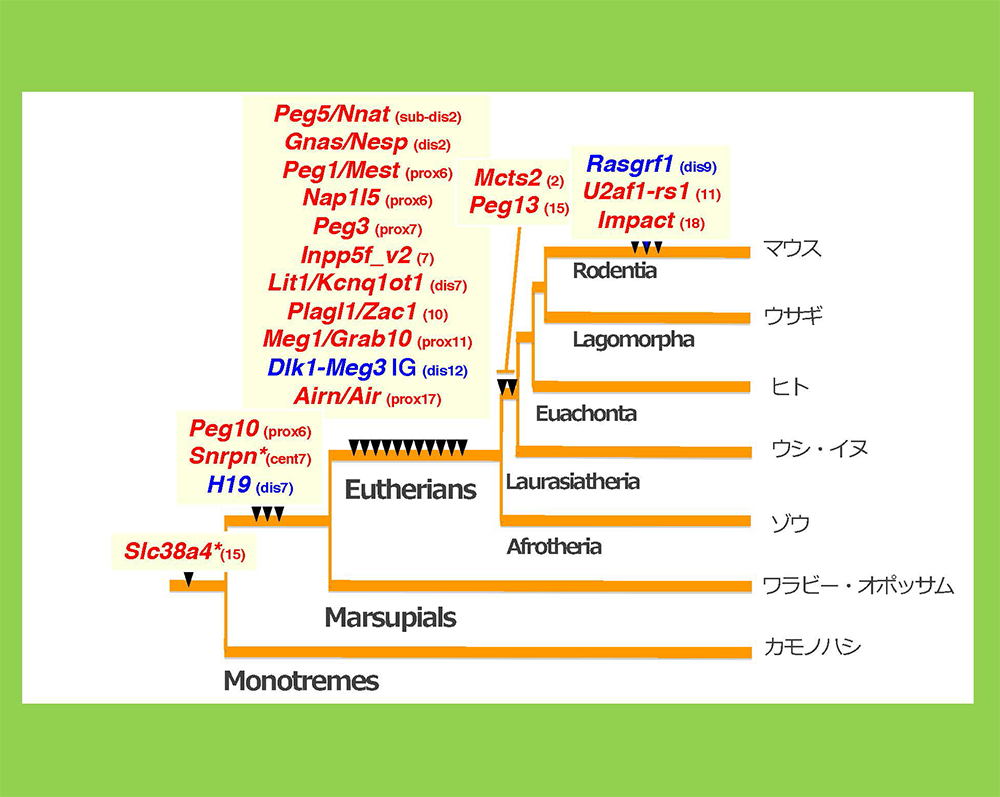
PEG10はレトロウイルス(LTRレトロトランスポゾン)のGAGとPOLに由来する獲得遺伝子です(図2)。胎生の哺乳類であるヒトやマウスを含む真獣類とカンガルーやコアラなどの有袋類だけに存在し(小野ら Genomics 2001, 鈴木ら PLoS Genet 2007)、胎盤形成に必須な機能を果たしています(小野ら Nat Genet 2006)(ヘッドライン4)。
胎盤で重要な機能を果たす遺伝子に、同じくゲノムインプリンティング研究から見つかったPEG11/RTL1があります(Charlierら Genome Res 2001)。これもGAGとPOLに由来する獲得遺伝子で、真獣類だけに存在します(図2)。PEG11/RTL1には真獣類の胎盤において母親からの栄養や酸素供給に重要な働きをする 胎児毛細血管 を維持する機能があり(関田ら Nat Genet 2006、鏡ら Nat Genet 2006)、これの獲得が真獣類の長期間の妊娠期間を可能にしたと考えられます(ヘッドライン5)。
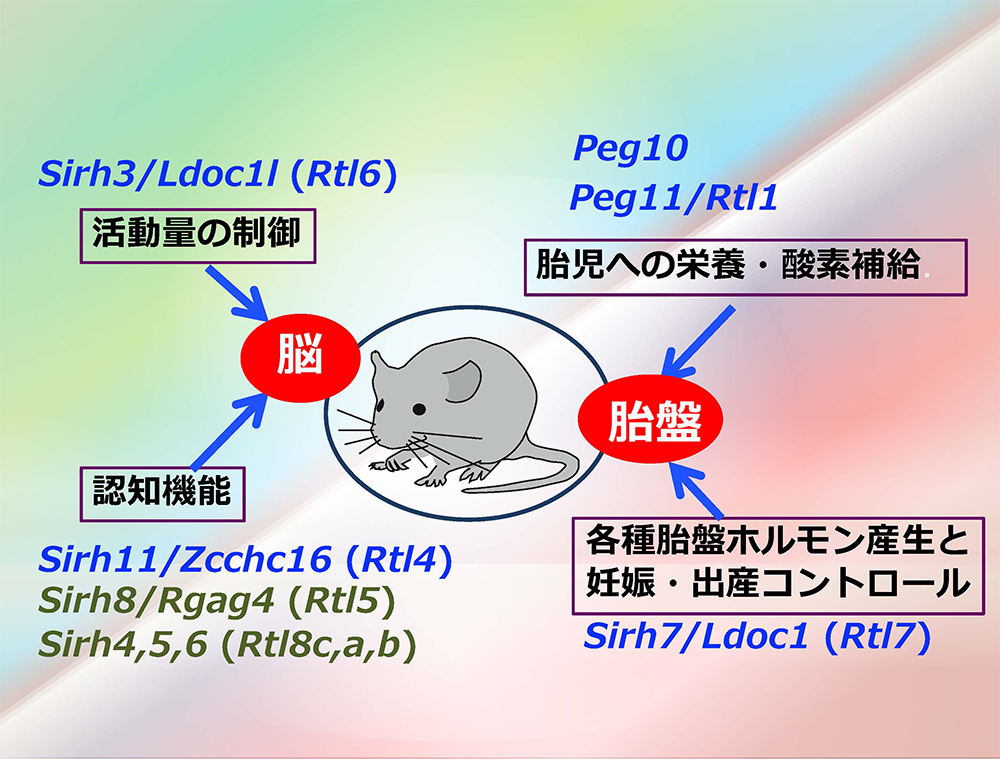
ヒトやマウスを含む真獣類には、PEG10, PEG11に加えて、その仲間の哺乳類特異的な獲得遺伝子が他に9個存在しています。PEG10, PEG11と同じレトロウイルス(LTRレトロトランスポゾン)のGAGから由来したもので、私たちはこれをSIRH/RTL遺伝子群と呼んでいます(図2)。
胎盤では、この他にSirh7/Ldoc1/Rtl7が機能しています。Sirh7は内分泌機能に関わることで出産タイミングの決定に重要な役割を果たしており、産子の生存に大きく関わる必須の遺伝子です(成瀬ら Development 2014)(ヘッドライン5、スピンオフ1)。さらにPEG10は、初期胎盤形成に加え胎児毛細血管の維持の働きもあり、妊娠期間全体で重要な働きをしていることもわかってきました(志浦ら Development 2021)(ヘッドライン10)
脳機能に関してSIRH/RTL遺伝子群は、神経細胞(ニューロン)とミクログリアで機能する2群に分かれます。
神経細胞(ニューロン)で機能するものに、GAG由来のSirh4, 5, 6/Rtl8c, a, bがあります(藤岡ら Open Biol 2025)。これらは3つの相同遺伝子がX染色体上にクラスターをつくっていますが、そのうち2つを欠失させた(Sirh5, 6/Rtl8a,b DKO)マウスでは成体期からの肥満、社交性の低下、無気力の亢進、子育て行動異常などの異常行動が見られました。その表現型の類似からゲノムインプリンティング疾患のPrader-Willi症候群の原因遺伝子である可能性が示唆されています(ヘッドライン8)。
また、「胎盤を創る」で登場したPeg11/Rtl1は、マウスではその欠失や発現過剰が胎盤の異常に加えて、運動性機能の低下、不安様行動の増加を引き起こし、加えて発現過剰では空間記憶の著しい低下も引き起こします。ヒトでもPEG11/RTL1の欠失がゲノムインプリンティング疾患のTemple症候群、発現過剰がKagami-Ogata症候群の原因となっています(ヘッドライン7)。
Peg10も脳特異的conditional KO(脳だけでPeg10を欠失させた)マウスで、脳機能への重要な関与が明らかになっています(志浦 未発表データ)。また、脳の神経細胞でのPEG10タンパクの増加がゲノムインプリンティング疾患であるAngelman症候群や筋萎縮性側索硬化症(ALS)で報告され、これらの疾患の原因となっている可能性が示唆されています(Pandyaら Cell Rep Med 2021, Whitelayら J Biol Chem 2021, Blackら eLife 2023)。
Sirh3/Rtl6、Sirh8/Rtl5、Sirh10/Rtl9、Sirh11/Rtl4/Zcchc16の4つの遺伝子は脳内の免疫細胞であるミクログリアで働く遺伝子でした。Sirh11以外の3つは病原体感染から脳を守る働きに関与しています(入江ら Development 2022、石野ら Int J Mol Sci 2023)。
自然免疫は細菌類、ウイルス、カビなどの真菌類の感染から私たちを守ってくれる最初の防御ラインです。これは広く動物界に保存されている進化的に古い機構です。各種のToll-like receptors (TLRs)が、細菌の細胞壁のリポポリサッカライド(LPS)、ウイルスのdsRNAや非メチル化DNAそしてカビの細胞壁であるザイモザンなどのpathogen-associated molecular patterns (PAMPs)を認識し、炎症反応を誘起して対抗します。脳は他の臓器と違い、ミクログリアが唯一の免疫細胞として自然免疫に関わります。SIRH3、SIRH8タンパク質はミクログリアから分泌され、脳内に広く分布していますが、それぞれが認識するPAMPsを検知すると、素早く複合体を形成して拡散を防ぐとともに、速やかな排除につなげます。一方、SIRH10タンパク質はミクログリアのリソゾームに局在し、取り込まれたザイモザンの分解に関わります。獲得遺伝子がPAMPsの排除に関わるという事実の発見から、真獣類では脳の自然免疫機構に特有の進化が起きていることが明らかになりました(ヘッドライン9)。
一方で、Sirh11/Rtl4/Zcchc16は別の機能をもっています。私たちは以前に、Sirh11ノックアウトマウスが新規環境への適応不全、衝動性の亢進、短期空間記憶力の低下を引き起こすことを報告しました(入江らPLoS Genet 2015)。ヒトではこの遺伝子の変異が自閉症スペクトラム症の発症に関わる(Limら Neuron 2013)ことから、この遺伝子から作られるSIRH11タンパク質が脳内で、どのような動態をするのかを解析しました。その結果、これがミクログリアから分泌され、覚醒状態では安定して存在すること、そしてストレスによって脳内に放出されるノルアドレナリンという神経伝達物質に応答することがわかりました(石野ら Int J Mol Sci 2024)(ヘッドライン9)これも、進化的には非常に古くて重要なホルモンであるノルアドレナリンを、より安全に効率よく脳で働かせるための、真獣類における進化であると考えられます。
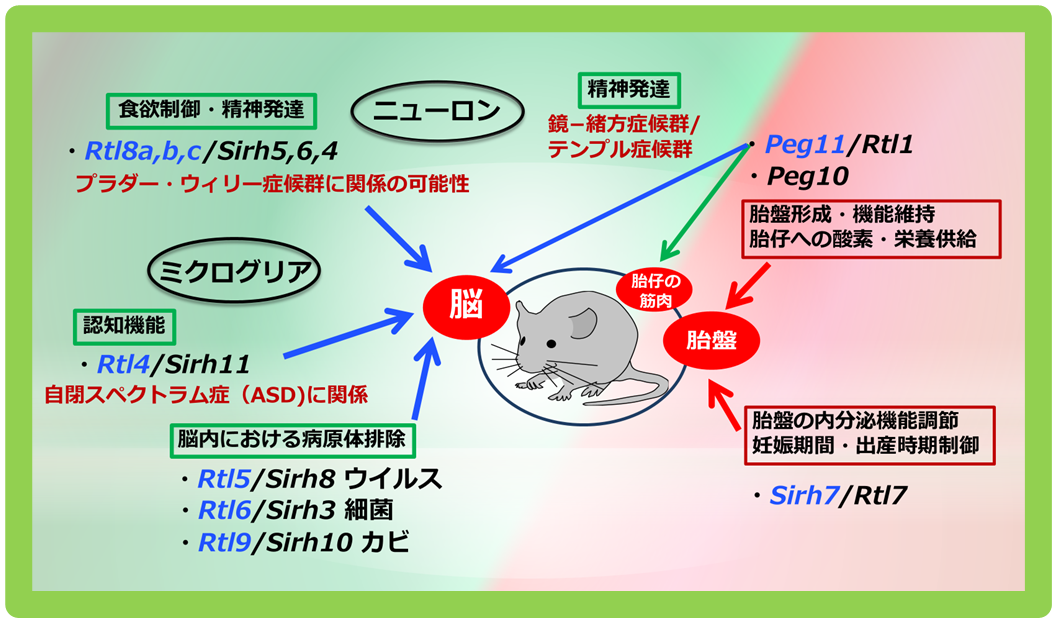
11個のSIRH/RTL遺伝子のうち、機能解明された10遺伝子の働きをまとめると図3になります。胎盤と脳という哺乳類を特徴づける2つの臓器でウイルス由来の獲得遺伝子が機能するという全体像が見えてきました
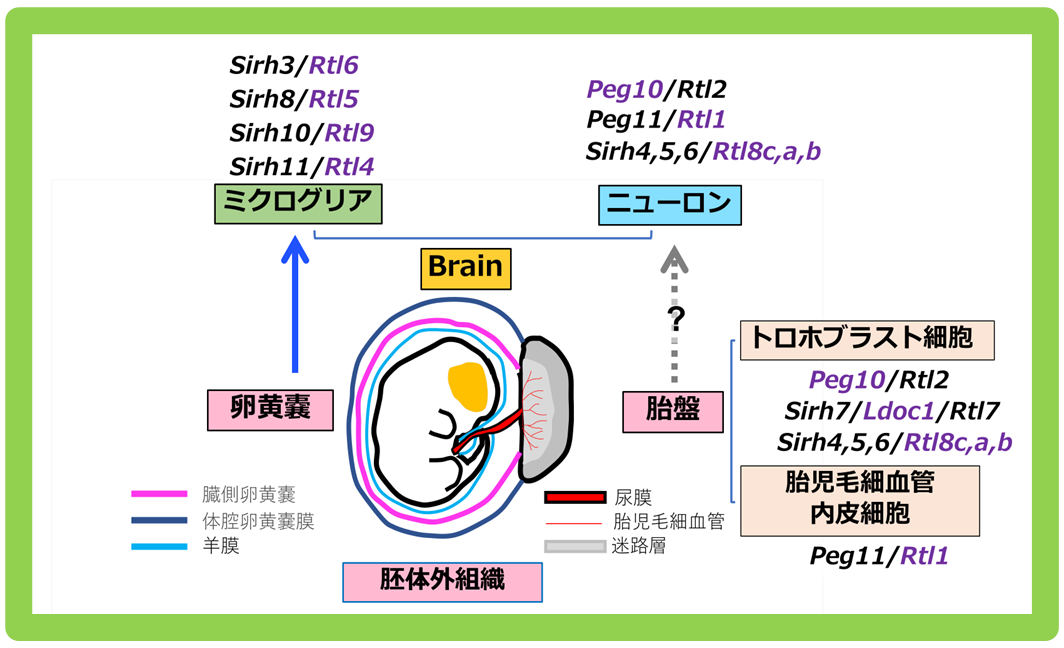
胎盤と脳はかけ離れた臓器のようですが、ミクログリアは個体発生の初期に卵黄嚢で発生し、胎児の脳に移動した細胞ですから、胎盤と同じ胚体外組織(胎児以外の組織)由来という共通点があります。このことは、これらの遺伝子が獲得された場である胎盤や卵黄嚢という胚体外組織の重要性を示していると考え、私たちは(以下の)新しい仮説を提唱しています。「胎盤や卵黄嚢などの胚体外組織は、遺伝子の発現を抑える働きがあるDNAのメチル化が胎仔と比べて低く、ゲノムに飛び込んだウイルス遺伝子を完全には抑え込まない。そのため胚体外組織は、変異によってウイルス由来の遺伝子に有利な機能が生じた場合に、新しい遺伝子として選択し、獲得する場として機能する。言わば進化の実験場として機能し、真獣類の進化を加速した」というものです。(図4)。よって、まず胎盤で機能する遺伝子や卵黄嚢を起源とする遺伝子が獲得されることになります。さらに胎盤のために獲得した遺伝子Peg10, Peg11などは、ニューロンでの機能も獲得しています。どのようにニューロンでの機能を獲得したのか興味が持たれますが、胚体外組織で獲得した遺伝子は、かたや胎盤を介してニューロンから、かたやミクログリアからと、2方向から脳機能へ関与することで、真獣類における脳の進化を加速した可能性も示唆しています。ヘッドライン10で詳しく紹介しますが、真獣類の胎盤形成には少なくとも5つのウイルス由来の遺伝子が関与しています。新たに誕生した胎盤は、さらに新しい遺伝子の獲得の場として機能し、真獣類の進化を促進するのに貢献したと考えています。
SIRH/RTL遺伝子群は外来(ウイルス由来)のDNAが、哺乳類の祖先が生きていた時代にゲノムに入り込み、変異により新規の機能を持ち、進化の過程でその有益性が自然選択された結果、哺乳類の新しい遺伝子となったと考えられます。
現在の“必然”としての日常は、“偶然”飛び込んできた外来のDNAが有益な遺伝子として選択されることで生み出されたといえるでしょう。
私たちの研究は、ウイルス由来の獲得遺伝子が哺乳類という大きなレベルの進化に寄与した具体例を示したものであると考えています。
ゲノムインプリンティングは動物では哺乳類だけがもっている遺伝子発現のしくみである。1984年に、片親由来のゲノムのみでは正常に発生しないことから、父親・母親由来のゲノムに差異があることが・・・
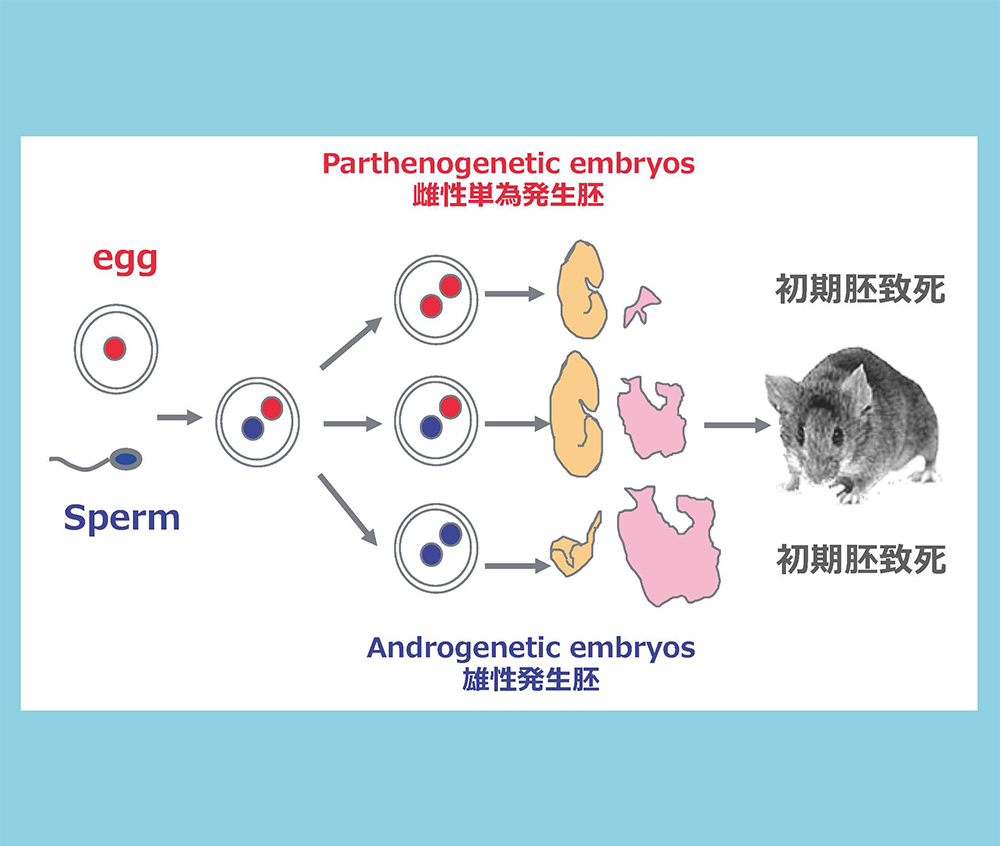
雌性単為発生胚と正常受精胚で発現に差のある遺伝子を引き算で濃縮する方法を開発し、父性発現遺伝子群(PEGと命名)の同定を進めた。この方法を主として12個のPEGを単離し・・・
インプリント遺伝子の発見に伴って、ゲノムインプリンティングは領域制御の片親性遺伝子発現機構であり、その制御の中心的な役割を果たすのは、雌雄のゲノムでDNAメチル化の差異を示す領域・・・
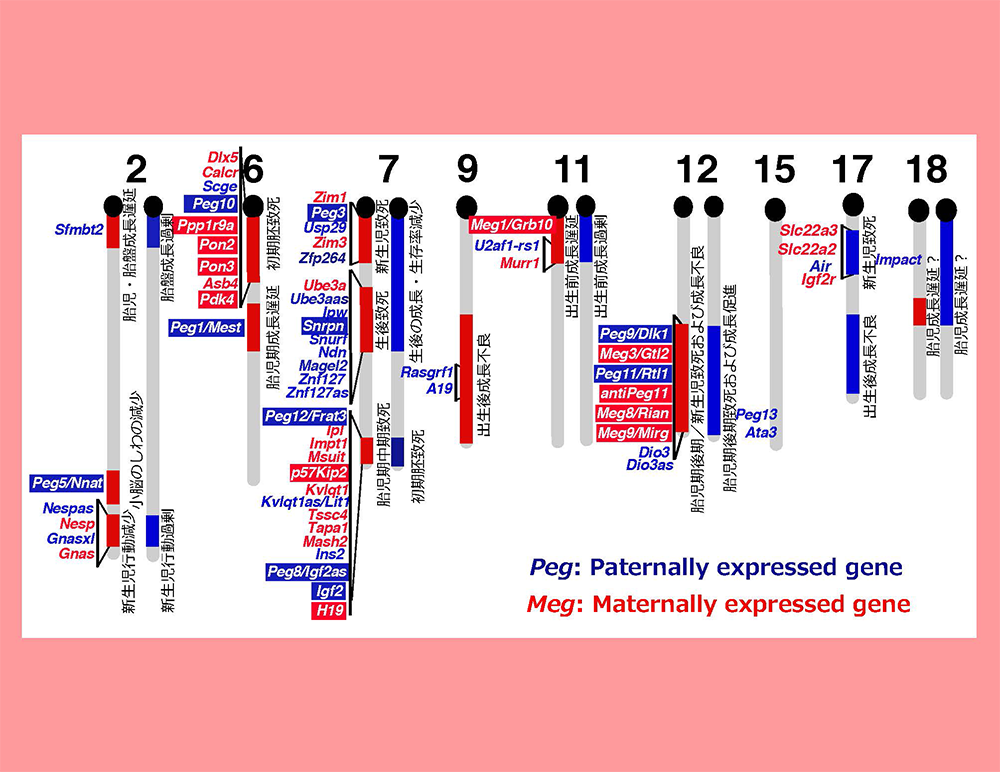
胎盤形成に必須なインプリント遺伝子は、母親性2倍体が初期胚致死を起こすマウス6番染色体近位部のインプリント領域にあると考え、ヒト相同領域の探索からPEG10遺伝子を発見した。
マウス染色体12番遠位部のインプリント領域では母親性2倍体マウスが胎児期後期からの成長遅滞や胎児期後期・新生児致死という表現型が見られる。我々はこれが胎盤の機能異常によるもの・・・
なぜ、哺乳類では発生に重要な遺伝子が片親性発現という生存に不利な機構が維持されているのか? 両親由来の2本のゲノムから遺伝子発現をすることは、劣性遺伝病の発症を防ぐ機構として・・・
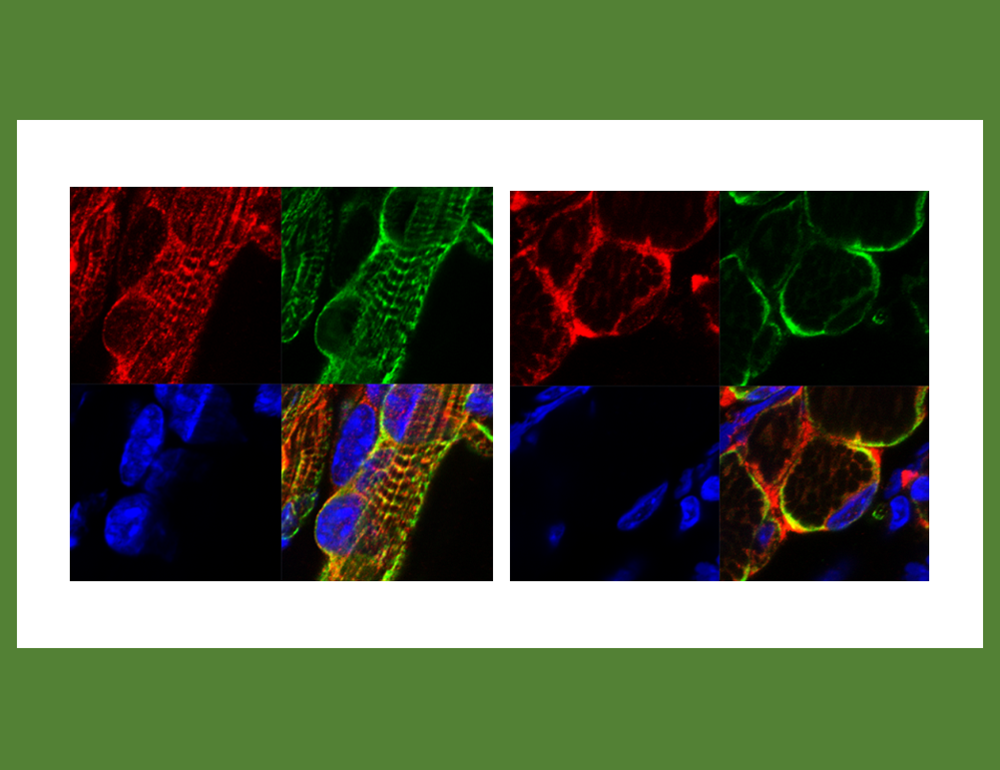
哺乳類らしさの構築にPEG11/RTL1が重要な貢献した2つの新しい証拠が加わった。胎児期及び新生児期特異的に筋肉発生に関わること、脳神経系の発達にも重要な機能を持つこと・・・
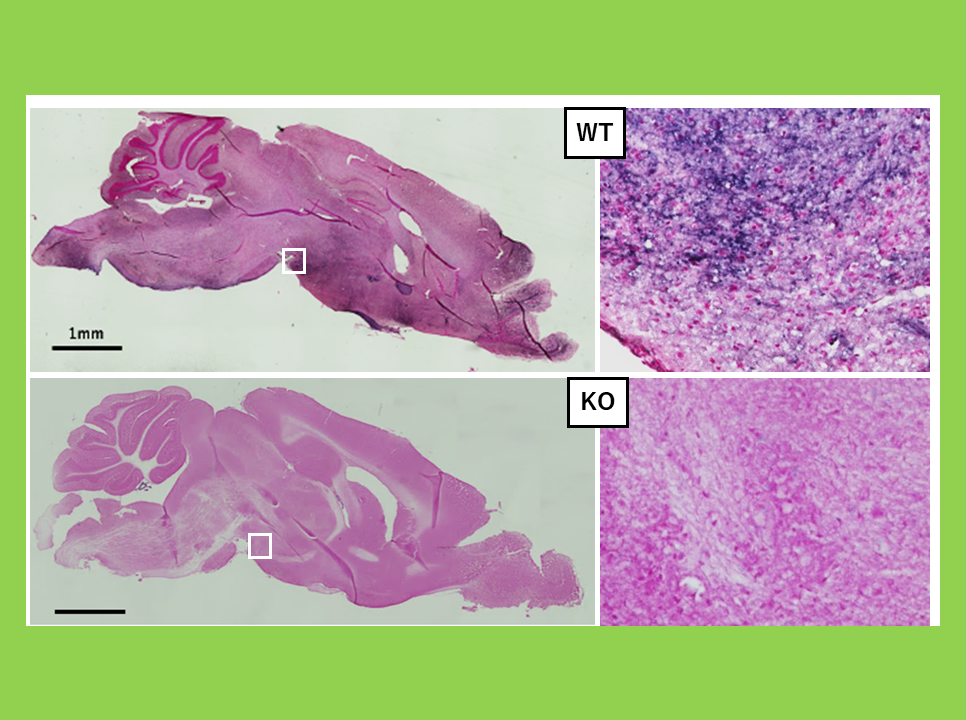
ニューロンで機能する遺伝子としてヘッドライン7で紹介したKagami-Ogata症候群、Temple症候群の原因遺伝子になっているPEG11がある。最近、PEG10もニューロンで発現し、・・・
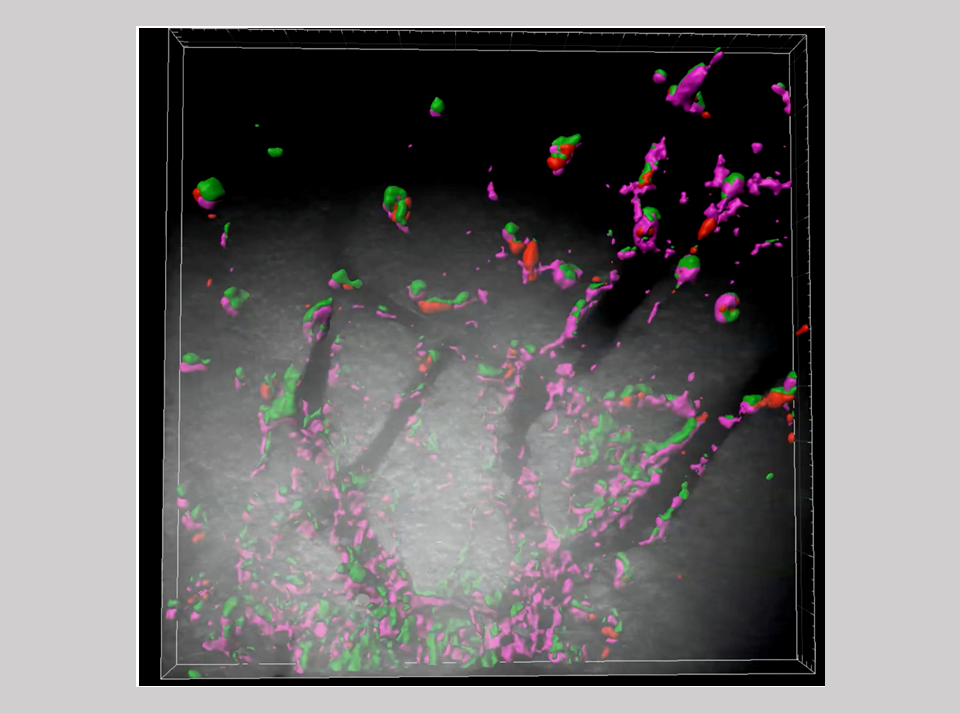
自然免疫は、動物界に広く保存される基本的な生体防御システムで、病原体など体に異物が侵入した時、マクロファージや好中球など貪食作用を持つ細胞がいち早く反応し排除する働きをしている。・・・
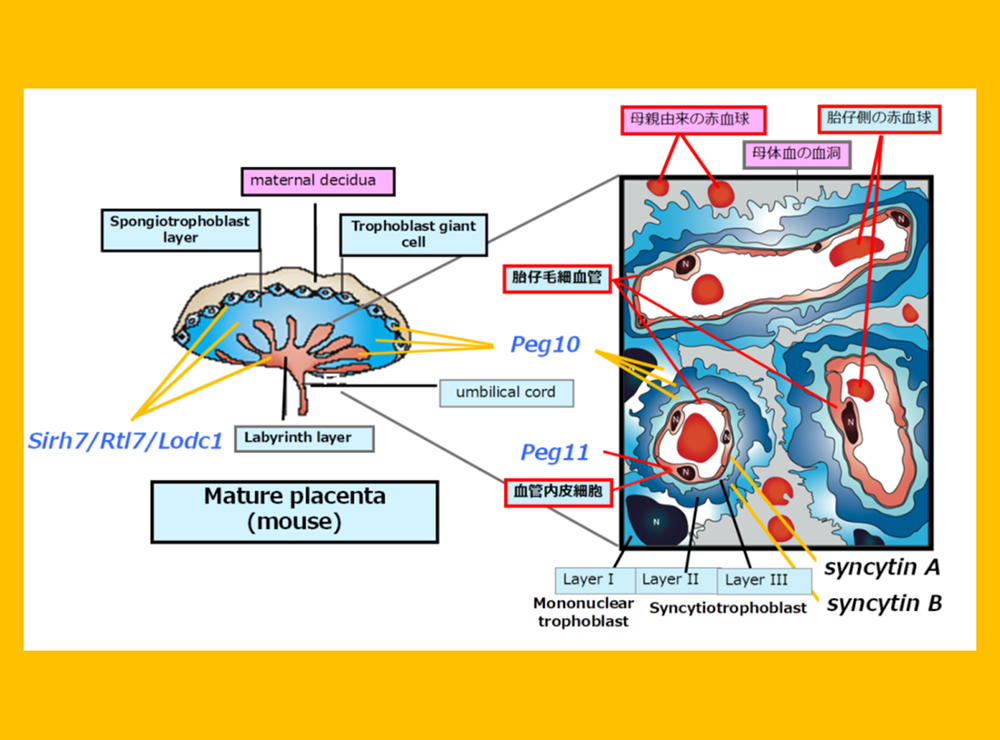
ここまで、PEG10が胎盤の初期形成に(ヘッドライン4)、PEG11/RTL1が胎盤内の胎児毛細血管の維持に(ヘッドライン5)、そしてSirh7/Ldoc1が胎盤の内分泌制御に機能(ヘッドライン5)・・・